ПОКАЗАТЬ ПЛАНПЕРЕЙТИ В КОНЕЦ
Лекция 2
Биоэнергетические функции митохондрий
Нарушение функций митохондрий при тканевой гипоксии
Нарушение биоэнергетических функций митохондрий - одно из наиболее ранних проявлений повреждения клеток.
Биоэнергетические функции митохондрий
Митохондрии - это везикулярные структуры, образуемые наружной и внутренней мембранами (см. рис.1). Внутренняя мембрана образует складки, или кристы, окружающие матрикс. На складках внутренней мембраны видны грибовидные выросты - это H+-АТФаза, или АТФ-синтаза, о которой будет сказано позже.
Внутренняя мембрана митохондрий непроницаема для большинства ионов, включая ионы водорода, натрия, калия, хлора. Но она, как и другие биологические мембраны проницаема для воды; поэтому объем митохондрий зависит от концентрации ионов внутри и снаружи. В норме концентрации частиц по обе стороны митохондриальной мембраны равны, при патологии концентрация внутри обычно повышается и митохондрии "набухают", к чему мы ещё вернемся.
Окислительное фосфорилирование
Митохондрии осуществляют важнейшую для клеточной биоэнергетики реакцию фосфорилирования АДФ с образованием АТФ за счет энергии окисления органических соединений, служащих субстратами окисления, молекулярным кислородом. Конечная стадия этого процесса - перенос электронов от восстановленных пиридиннуклеотидов и сукцината на молекулярный кислород осуществляется по системе переносчиков электрона, которая в совокупности называется дыхательной цепью (см. рис.2).
При переносе электронов по дыхательной цепи происходит высвобождение энергии, величина которой (в электрон-вольтах) равна разности стандартных восстановительных потенциалов двух реагирующих ред-окс пар. Эти стандартные потенциалы приведены на рис.3.
Энергия одного моля иона в данной среде называется, как известно, электрохимическим потенциалом. Разность электрохимических потенциалов протона между двумя водными фазами внутри и вне митохондрий описывается уравнением:
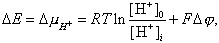 |
(1) |
где R - газовая постоянная, T - абсолютная температура, [H+]o и [H+]i - концентрации ионов водорода вне и внутри матрикса, соответственно, F - число Фарадея, Dj - разность потенциалов между окружающей средой и матриксом.
Петер Митчелл в качестве единицы энергии использовал электрон-вольты, в результате чего уравнение (1) несколько трансформируется:
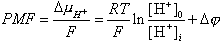 |
(1) |
Суммарная энергия окислительно-восстановительной реакции, превращенная в разность электрохимических потенциалов ионов водорода, была названа П. Митчеллом протон-движущей силой (PMF - proton motive force), по аналогии с электродвижущей силой в гальванической батарее. Заменив натуральный логарифм десятичным, легко найти величину протон-движущей силы, зная разность pH (DpH) и разность потенциалов (Dj) между средой и матриксом при комнатной температуре; выраженная в милливольтах она будет равна:
| PMF (мВ) = 60 (мВ) · DpH + Dj |
(2) |
В митохондриях основной вклад в эту сумму вносит мембранный потенциал, который в присутствии субстрата и кислорода составляет около 170-180 мВ.
Созданная работой дыхательной цепи разность потенциалов DmH+ может быть использована для синтеза АТФ или переноса ионов в митохондрии. Синтез АТФ осуществляется благодаря работе АТФ-синтазы, которая представляет собой протонную АТФазу (H+-АТФазу), работающую как бы "задним ходом" (см. схему на рис.4).
Перенос ионов через мембрану митохондрий
Альтернативный путь использования накопленной в форме DmH+ энергии - транспорт ионов внутрь митохондрий. Рис.5.
Поскольку внутри митохондрий при энергизации появляется отрицательный потенциал, катионы засасываются электрическим полем в матрикс, если мембрана для них проницаема. Во внутренней мембране имеется переносчик ионов кальция. Движение кальция снимает мембранный потенциал (Dj в уравнении 2), что приводит к довольно резкому возрастанию DpH, т.е. защелачиванию матрикса (см. рис.6).
С другой стороны, между матриксом и окружающей средой в энергизованной митохондрии имеется разность pH (более щелочная среда внутри). Это приводит к диффузии внутрь митохондрий ортофосфата, для которого в мембране имеется переносчик, работающий по механизму антипорта: ион HPO3- обменивается на гидроксил OH- (рис.7). В водном растворе это эквивалентно симпорту (совместному переносу) H+ и HPO3-.
При таком переносе не происходит переноса заряда через мембрану, и поэтому перенос не зависит от мембранного потенциала. Однако разность концентрации H+ способствует переносу фосфата в матрикс по закону диффузии для ионов гидроксила, которых в матриксе больше, чем снаружи.
Таким образом, ионы кальция накапливаются в матриксе электрофоретически, при этом мембранный потенциал снимается, а разность pH на мембране возрастает. Фосфат переносится в матрикс в конечном счете в электронейтральной форме, при этом снижается разность pH, а мембранный потенциал растет. Если в среде присутствуют одновременно и ионы кальция и ортофосфат, происходит накопление обоих в матриксе, при этом снимается как мембранный потенциал, так и DpH. Ничто более не препятствует свободному прохождению ионов водорода через мембрану в точках сопряжения, и дыхательная цепь работает на полную мощность.
Из теории Митчелла следует, что прекращение синтеза АТФ митохондриями (или подавление транспорта кальция и фосфата в матрикс) может произойти не только в результате повреждения дыхательной цепи (и снижения скорости дыхания, т.е. окисления субстратов кислородом), но и в случае нарушения барьерных свойств внутренней мембраны. В последнем случае происходит утечка ионов через поврежденную мембрану и падение разности потенциалов на мембране, необходимой для работы АТФ-синтазы, транспорта фосфата и ионов кальция.
Функциональные состояния митохондрий по Б. Чансу
Изучение функций митохондрий и их нарушения производится после выделения этих органелл из ткани; при этом важно не повредить митохондрии в ходе самого выделения.
Один из способов изучения функции митохондрий - измерение скорости потребления кислорода суспензией органелл в различных функциональных состояниях методом полярографии. На рис.8 приведена схематизированная запись изменения концентрации кислорода в суспензии в ходе инкубации изолированных митохондрий.
Наклон кривой в каждый момент времени характеризует скорость потребления кислорода (дыхания) в данном состоянии, эти величины принято обозначать как V1, V2, V3, V4 и т.д., где цифрами обозначены состояния по классификации Б. Чанса. Наиболее информативны V3 - скорость дыхания митохондрий при окислительном фосфорилировании, т.е. в присутствии субстратов окисления, АДФ и ортофосфата, и V4 - скорость дыхания митохондрий в присутствии субстратов окисления и ортофосфата, но в отсутствие АДФ (состояние дыхательного контроля).
На рисунках 9.1, 9.2 и 9.3 показано, что именно происходит в митохондриях в этих состояниях.
2-6 - основные функциональные состояния, U - разобщенное состояние митохондрий.
Рассмотрим вопрос подробнее.
- - это состояние митохондрий, к которым не добавлены ни субстраты, ни АДФ (- SH2, - АДФ). Состояние неопределенное, т.к. сколько-то эндогенных субстратов и АДФ есть в самих митохондриях;
- - к митохондриям добавили АДФ, но не добавили субстраты окисления (- SH2, + АДФ). Это деэнергизованное состояние, т. к. в отсутствие субстратов дыхательная цепь не работает и митохондрии не поддерживают мембранный потенциал (см. рис.9.1);
- - состояние окислительного фосфорилирования (+ SH2, + АДФ). Окисление субстратов приводит к энергизации митохондрий, мембранный потенциал тянет протоны внутрь через АТФ-синтазу и происходит синтез АТФ из добавленного АДФ (см. рис.9.1);
- - состояние дыхательного контроля (+ SH2, - АДФ): митохондрии энергизованы, но дыхание подавлено, поскольку в отсутствие АДФ высокий мембранный потенциал останавливает перенос протонов через мембрану и следовательно - перенос электронов по дыхательной цепи (см. рис.9.2);
- - состояние анаэробиоза (в среде кончился кислород). Митохондрии деэнергизованы и, разумеется, не фосфорилируют независимо от того, есть в среде субстраты и АДФ или нет (см. рис.9.2);
- - состояние ионного транспорта. При добавлении субстратов и ионов кальция последние втягиваются внутрь электрическим полем, снижая мембранный потенциал. Одновременно внутрь начинает закачиваться ортофосфат, под действием DpH. В итоге внутри накапливается фосфат кальция, а DmH+ снимается, и дыхательная цепь работает с максимально-возможной скоростью (см. рис.9.3);
U - к митохондриям добавлен переносчик протонов через мембраны (например, 2,4-динитрофенол). В результате движения протонов внутрь снимается DmH+ и дыхательная цепь работает вовсю (см. рис.9.3).
Вопросы для самоконтроля
- Общая схема строения митохондрий. Дыхательная цепь. Восстановительные потенциалы переносчиков. Точки сопряжения.
- Схема окислительного фосфорилирования по Митчеллу.
- Перенос ионов кальция и фосфата в митохондрии.
- Функциональные состояния митохондрий. Условия, скорость дыхания и мембранный потенциал в каждом из состояний.
- К каким изменениям в скорости дыхания приведут повреждения дыхательной цепи и нарушение барьерных свойств мембран?
ПОКАЗАТЬ ПЛАНПЕРЕЙТИ В НАЧАЛО